Maynard Smith & Szathmáry (1995) pointed out that the development of a multicellular organism, with differentiated cells, requires that three problems be solved. The first problem is gene regulation. Only very sophisticated mechanisms for gene regulation can produce differentiated cell types. This problem is not addressed in this thesis. The second problem, cell heredity, is irrelevant as far as the slime mould is concerned. During morphogenesis no cell division takes place, and hence states of differentiation do not have to be (partly) heritable through cell division. The third problem, which we have focused on, is the emergence of morphogenesis: how morphs can be generated and how a multicellular creature can sense and response in a coordinated way to the environment.
With the model for Dictyostelium discoideum presented in this thesis we have demonstrated that it is possible to study morphogenesis in the widest sense of the word, and to identify the underlying mechanisms. These mechanisms will be reviewed in this final chapter; next we look in more detail at the modelling approach we have used; and finally we point to some omissions in our model, as well as to its potential as a basis for interesting further research. First of all, however, the main themes of the thesis are briefly reviewed.
Part I of the thesis is devoted to pattern formation in excitable media. In chapter 2 we show the occurrence of lateral instability in excitable media, which leads either to propagating waves with ripples, or to spiral breakup. Lateral instability can develop when the inhibitor diffuses at a high rate, and is associated with a negative slope of the so-called eikonal-curvature relationship: when the inhibitor has a high diffusion rate, the velocity of a wave is restrained by the inhibitor ahead of the wavefront, and hence the wave moves faster when the wavefront is slightly curved. The slope of the eikonal-curvature relation could be measured using quasi one-dimensional computations, and is used as fitness measure in a genetic algorithm. In this way we were able to zoom in on the parameter region where lateral instability could be observed. We also found a number of other types of interesting dynamics, the most notable being a regime with diffusion-driven limit-cycle behaviour.
Our study, when combined with previous studies that used the same model (see, e.g. Panfilov & Hogeweg, 1996,1993), shows that the model we use is well suited for investigating the very wide range of dynamics that occur in excitable media.
Part II of the thesis is devoted to Dictyostelium discoideum development. The above-mentioned model is used to describe the cyclic AMP (cAMP) dynamics; a stochastic two-scale cellular automata (CA) model, presented in detail in section 1.3, is used to describe the individual amoebae. In the model, every cell is represented as a group of connected automata, i.e. the basic scale is subcellular.
Chapter 3 tackles problems connected with slug motion, such as shape conservation and cell sorting, as well as the mechanisms underlying thermotactic behaviour, which direct the slug to the surface (Dusenbery, 1988). We have implemented cell adhesion and cAMP signalling in such a way that they can interact, but it is still possible to unravel the mechanisms that emerge. We show that the interplay between differential adhesion and (non-differential) chemotaxis is sufficient to explain a number of aspects connected with normal slug motion, and also leads to fast and complete cell sorting, a phenomenon which cannot be due solely to cell adhesion or chemotaxis.
The whole slug is able to show an adequate response to a thermal gradient, by means of temperature-dependent differences in its excitability. Individual amoebae, on the other hand, can neither sense a temperature gradient, nor show temperature-dependent differentiation in motion velocity. The behaviour is achieved by a modification of the cAMP waves: differences in temperature alter the excitability of the cell, and thereby the shape of the cAMP wave. Chemotaxis towards cAMP causes the slug to turn. Furthermore, due to the fact that the wavefronts always keep a smooth shape because of the well-known curvature-effect, the waves of excitation function as spatiotemporal noise-filters. This spatiotemporal integration generates a very high sensitivity (even very small temperature gradients can be detected), as well as the ability to receive a signal in the presence of extreme noise.
In conclusion, the study demonstrates the importance of the interactions between different processes, and shows that precise sensing of the environment can result from the integration of spatiotemporal information.
Next, the model is extended with ammonia (NH3) dynamics and light refraction. In chapter 4 we show that the phototactic response, which leads the slug to the soil surface, can also unfold from the interactions between the different modules. Again the individual amoebae are unable to detect the signal (where does the light come from?), or to show an adequate response (move towards the light source). We have elucidated the following minimal mechanism for phototaxis: because the slug refracts the light and has a rounded shape, the light is focused on the side opposite the light source (the `lens-effect'). Production of NH3 increases with light intensity, and the NH3 decreases the excitability. Hence waves move more slowly on the side opposite to the light source, they become slanted, and chemotaxis towards the waves causes the slug to turn.
This study illustrates how side-effects1 can be important for validating the model, for generating new ideas, and for gathering together multiple aspects of the development: we did not set out to explain mutants which show bi-directional phototaxis or to interpret experiments that were done with these mutants, yet we are able to explain these as autonomous results (see also section 7.2).
Next, the model is extended with cell differentiation that stems from a local induction process, and with production and displacement of the extracellular matrices. We also introduce differences in stiffness. In chapter 5 and chapter 6 we move from the slug stage to the culmination stage. The `textbook-view' of the culmination has always been that the stalk grows by the successive addition of prestalk cells at the top, and the prespore cells are carried aloft by the growing stalk (Thomason et al., 1999). However, both our model and in vivo experiments performed by others rule out this possibility. We show that periodic upward movements, due to chemotactic motion towards cAMP, induce pressure waves, which squeeze the stalk downwards through the cell mass. These pressure waves result from cAMP dynamics and differential adhesion, i.e. they are themselves observables generated by the model. The mechanisms that we propose are in agreement with, and moreover, can explain many experimental observations. The mechanisms show a high amount of self-organisation and self-correction: an elongated stalk is formed which very rapidly moves downwards to the base; any deviation from downward elongation is corrected very efficiently; pathfinder cells always remain positioned along the tip of the stalk; the downward motion stops automatically when the stalk tip reaches the base and the same holds for the upward motion when the stalk formation is completed; a rounded spore head is formed on top of the stalk.
In conclusion, our study, when combined with the study by Savill & Hogeweg (1997), lets us describe within one model the entire process of morphogenesis from aggregation all the way to fruiting body formation (except for the initiation and termination of the slug stage). It can be said that our model of the culmination of Dictyostelium discoideum culminates the modelling of its development.
The two-scale stochastic CA model we used to describe the amoebae is ideal for studying the development of Dictyostelium, since the cooperative behaviour observed during the development is driven by the movements and responses of the individual cells. Besides, the formalism handles pressure, deformation and motion, i.e. the physical aspects of morphogenesis, in a very elegant way. Only local transition rules are involved in the description of these processes; there is, for example, no need to specifically calculate the global pressure distribution. In short, all update rules in the model are defined on two local scales, namely cellular and subcellular. Nevertheless, the behaviour that can be observed at the intermediate and global level is due to the collective behaviour of the cells: the multicellular organism that is formed by all the amoebae together can sense the environment much more precisely than an individual amoeba can, and can respond to it in a coordinated and accurate way, and surprisingly, even new observables emerge from the collective behaviour of the amoebae.
For example, an individual amoeba can neither respond to a slight temperature gradient, nor measure the direction of incidence of light. Both the very high sensitivity and the ability to filter the noise, as well as the ability to determine the light source, arise from the collective behaviour. The slime mould uses self-organising structures that are `larger than life', i.e. larger than the individual amoebae, on order to make information integration, information processing and information transmission possible. The shape of the slug itself behaves as a lens, waves of excitation are formed in which many amoebae are involved at every moment in time, broad pressure waves generate peristalsis, lumps of cells move with different velocities because of the interplay between chemotaxis and adhesion, and so forth.
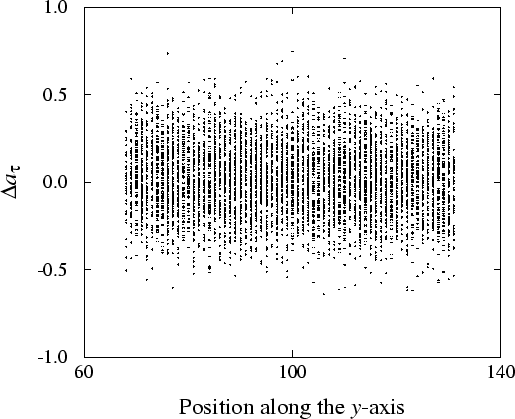
The mechanisms underlying thermotaxis and phototaxis are both related to slanted wavefronts. The chemotaxis then causes the slug to turn. Several properties of wave propagation play an important role. For example, in our Dictyostelium model the concave wavefronts move faster than plane waves, due to an increased influx of activator into the nearby region, and for the same reason convex wavefronts move more slowly (this is called the curvature-effect). It can easily be seen that due to this curvature-effect the short-wavelength noise can be averaged. This is why even very small temperature gradients can be measured and why even very high noise levels can be filtered: for example, the noise can be even more than 300 times as large as the signal (see Fig. 7.1).
 |
The curvature-effect also explains why the wave becomes slanted when during phototaxis there are large differences between the concentrations of NH3 on each side of the slug. When the slant becomes too large, the momentum of the force, created by the cells moving towards the cAMP wave, causes the side-effect of bi-directional phototaxis.
During culmination many self-organising and self-correcting phenomena occur, all of which are needed for successful fruiting body formation. A nice `side-effect', however, is the orientation of nearby culminants: the default production of NH3 alters the excitability, and therefore the waves again become slanted. As a consequence, the fruiting bodies end up orientated away from each other. This is often observed in experiments; whether or not this behaviour is functional, however, is still under debate.
In general, theoreticians show a preference for keeping models as simple as possible. And for good reason. Nevertheless, the model we end up with is rather complex by theoretical standards, although much of the complexity of Dictyostelium is still ignored (e.g. there is no gene regulation in the model).
In this section we first re-emphasise the need to study the simplest models; then we discuss why we think we have struck a useful balance in our model; and finally we discuss ways in which we can extend our modelling approach so as to incorporate gene regulation, while still pursuing our aim of uncovering general principles.
The usefulness of the simplest possible models is indisputable. First of all, one can understand the model thoroughly. Secondly, many aspects of a system under study, do not (or barely) influence specific phenomena of interest, and should be ignored. This allows us to consider models which are valid for many systems. A good example are the models of excitable media we discussed earlier; the concept of excitable media is in fact an abstraction which ignores irrelevant details. One should, however, bear in mind that, because of the generality of such representations, and because they are often used to study phenomena that occur generally, these models do not reveal details about the particular implementation. Thirdly, it is very useful to derive measures that are independent of much of the observed variation. A specific example that we would like to mention here demonstrates the derivation of such a measure for a particular case, namely the fitness of viral strains.
Fitness differences between viral variants are often estimated experimentally by means of pairwise competition experiments. As a fitness measure, one conventionally uses the slope of the logarithmic ratio of the genotype- or phenotype-frequencies plotted in time. This fitness measure, however, remains proportional to the actual replication rate realized in the particular experimental setup, and therefore cannot be extrapolated to other situations. We have developed a method of computing the relative fitness, which is independent of the actual replication rate (Marée et al., 2000, not part of this thesis). By using generic competition models, we were able to derive a formula for the relative fitness, without having to make any assumptions about (changes in) the replication rate during the course of the experiment. This means that we obtained a general formula that is applicable not only when viruses grow exponentially, but also for (quasi) steady state conditions, as well as for any other growth regime. A tool for calculating the relative fitness from observed changes in viral load and genotype (or phenotype) frequencies is publically available on our web-page.
Obviously, models should be simple, but not too simple. Recently we have developed a heuristic method which can help modellers to find sensible ways of extending very basic ordinary differential equation (ODE) models, in order to fit the data, yet can enable them to keep their models as simple as possible (Müller, Marée & de Boer, 2000b, not part of this thesis). In the latter study we first state that every parameter describes many underlying processes and that the value of a parameter in equilibrium is just a realization of the process function. Next we calculate parameter values from observed steady states, and replace the parameters with the highest variation by a more explicit description of the underlying processes. We have applied this method to HIV-1 set-points and obtained a model in which several factors collectively determine the steady-state viral burden. The model is unlike most ODE models, in which steady states of variables are usually determined by one process only. Our method gave a strong indication that the lymphoid tissue could play an important role in the viral dynamics. We have used this observation to extend the model further with such a compartment (Müller, Marée & de Boer, 2000a, not part of this thesis). Note, however, that this method can only be applied in systems in which the dynamics always reach a steady state. This imposes a strong limitation on the systems that can be studied, e.g. it precludes pattern formation.
In part II of this thesis we address a different level of model complexity. We use a comprehensive model, in which we do not confine beforehand the set of phenomena we look at; instead we implement a number of experimentally established processes that occur during the morphogenesis of Dictyostelium, and we observe the consequences of their interactions. We implement these processes using different model-formalisms. They were chosen on the basis of what they could describe in the easiest way, which helped to maintain a certain level of simplicity in the model, and allowed us to filter out everything which does not influence the phenomena of interest. For example, in our hybrid model light was described in a CA, using a ray-tracing technique, whereas NH3 was described in a partial differential equation, as a reaction diffusion equation; the tube mass could be very well described as one very large and stiff cell in the Glazier & Graner-model formalism. We have shown how many aspects of the morphogenesis emerge from the dynamic interactions on different scales between the generic behaviours of these different modules. Nowadays, several other research groups are also moving into this direction, and have also started to model phenomena `beyond pattern formation' (see, e.g. Bretschneider, Vasiev & Weijer, 1999; Murray & Swanson, 1999; Kerszberg & Wolpert, 1998).
Whereas we emphasise the importance of the interactions between different processes, many studies have instead tried to explain a specific feature with one linear mechanism. For example, Clow et al. (2000) have tried to ascertain whether cell sorting in Dictyostelium is due to differential cell adhesion or chemotaxis. First they showed that there is a massive evidence in literature that both processes are important; next, however, they tried to find out which idea must be wrong. They concluded that differential adhesion could be ruled out, because they found that clumps of cells did not slowly increase and fuse; instead they observed directed motion, which did not agree with their intuitive idea about what kind of patterns were to be expected if differential adhesion were the only process at work. However, they could not explain the patterns with chemotaxis towards cAMP alone either, and therefore postulated that another process precedes chemotaxis towards cAMP. However, if we assume that both chemotaxis and differential adhesion are at work, their observations fit quite well to our picture of cell sorting. Having identified the first cells that show the prestalk cell-specific marker as the most excitable ones and therefore the ones that become the organising centre of cAMP signalling, we expect to observe directed cell motion towards this initial cluster if there is differential adhesion between the cell types. We know that this initial cell sorting changes the properties of the excitable medium itself; these changes strongly couple back to the cell dynamics. The model we have developed can be used as a tool to analyse such complex interactions and feedbacks and to link them to the observations in the above-mentioned experiments.
As indicated at the beginning of this section, we have completely ignored gene regulation in our model. We find that the whole development of Dictyostelium can unfold without the need to change the parameters of most processes: in our model the morphogenesis is mainly a self-organising process, and many interesting new observables emerge solely from the interactions between generic modules. This view, however, deviates from the general genetic way of looking at development in terms of gene regulation.
Although Dictyostelium is considered to be the most primitive eukaryote which has the basic features of multicellular development, it still has 8000 - 10,000 genes, of which, for example, at least 3000 are expressed during the slug stage (Loomis, 2000). The rapid increase in genetic knowledge challenges us to determine functional roles for all those genes. We are aware that many of these genes are involved in adhesion, induction, maturation, sensing of the environment, etc.; in our opinion, for a successful interpretation of the role of these genes, we need to understand the dynamics and mechanisms on the intermediate scale.
In this thesis we opted for a level of abstraction at which we did not implement the specific function of individual genes. Many models, however, have been developed to explain how the generic principles that we have used are implemented at the (sub)cellular level (although so far few models incorporate the level of the gene regulation). Interpreting gene expression in terms of our model presents a challenge; in particular, it may well prove possible to link distortions of (sub)cellular processes to aberrant phenotypes.
The technique of restriction enzyme-mediated integration (REMI) makes it possible to create many aberrant phenotypes (Smith, 2000). REMI allows both the production of mutants and the simultaneous tagging of the mutant gene. Flanking genomic sequences of significant mutants can be recovered, cloned and sequenced (Newell & Gross, 2000). Thus, information can be obtained about both the aberrant morphogenetic process and the gene involved, e.g. by searching for homologues. When the role of a gene at the (sub)cellular level can be elucidated, and its effect on the module can be determined, our modelling approach should be able to predict and explain the aberrations that can be observed during the morphogenesis. In conclusion, interpreting gene expression in terms of our model provides a heuristic method for assigning gene function in the light of the morphogenesis as a whole.
Recent studies have suggested that organisms may need complex gene networks in order to function robustly. Barkai & Leibler (1997) and Alon et al. (1999) have shown that in the biochemical network which organises chemotaxis in the bacteria Escherichia coli, the precision of adaptation is robust. In contrast, other properties, such as steady state behaviour, are less robust. They found that the precision of adaptation is a consequence of the network's connectivity and does not require `fine-tuning' of the biochemical parameters. von Dassow et al. (2000) looked at the gene network which fixates the segment polarity pattern in the embryo of the fruitfly Drosophila, and also found the network to be robust, in the sense that correct patterning is relatively often found when parameters are varied randomly.
The question that then arises directly is whether our model also needs a complex gene network to ensure its robustness. Since the modules we use are very coarse descriptions of the underlying dynamics, our model behaviour is in some sense very robust with respect to particular implementations of the (sub)cellular processes. However, we had to zoom in on a very specific parameter space to find the phenomena we were seeking. Thus, the specific morphogenesis presented here is not a generic property of the model in the way that the spiral wave is a generic property of many models that describe excitable media. Nevertheless, once we found the right parameter region, it turned out that we could always vary parameters considerably, yet retain the model's qualitative behaviour; we have found many self-organising and self-correcting properties, as a result of which the observed phenomena are not too sensitive to the specific parameter values. Nevertheless, because certain processes have to remain within this (relatively broad) range of admissible parameter settings, it is particularly the adaptation to various circumstances which has to be robust (cf. Alon et al., 1999).
The previous section also raises the question of how such gene networks, connected to development, evolve. Indeed we started this chapter by stating that the development of a multicellular organism requires that besides the problems related to pattern formation, problems related to gene regulation and cell heredity have to be solved as well (Maynard Smith & Szathmáry, 1995). Evolutionary processes have not been taken into account in this thesis. Nevertheless, it is interesting to speculate on how these questions might be incorporated in our modelling approach.
Hogeweg (2000b,a) has taken up this challenge. She has made precisely the above-mentioned extensions to describe all the elements of the evolution of multicellularity (as indicated at the beginning of this chapter), while at the same time minimising the a priori assumptions. The problem tackled in these studies is multicellular development in general; no specific organism has been modelled. The first new element, i.e. gene regulation, was implemented as a boolean network defining genes that encode cell signalling, maternal factors and adhesion strengths. The second new element, i.e. cell heredity, was implemented by allowing for cell divisions during early development as well as when cell volumes are large. After cell division both daughter cells keep the same state of the network, i.e. there is cell heredity2. The gene networks can evolve; the only fitness measure that is used is simply cell differentiation. Now, with all four basic elements of the evolution of multicellularity implemented at (sub)cellular level, but with no direct solutions included (and with even fewer parameters than in our Dictyostelium model), we see the emergence, in many evolutionary runs, of cell differentiation, gene-regulation and a balance between cell heredity and positional signalling, as well as, most surprisingly, many types of morphogenesis: in the `shadow' of the evolutionary run phenomena are observed which could be best described as engulfing, budding, elongation, intercalation, meristematic growth, and so forth. This means that this kind of modelling is able to generate morphogenesis, solely as a side-effect of the need for cell differentiation during the transition to multicellularity.
Research into systems in which the behaviour can be easily understood has made it simpler to focus on `generic' properties occurring in many contexts. The above-mentioned study illustrates that it seems profitable not only to study generic properties of minimally defined systems, but also to derive general properties of relatively `rare' and complicated systems while minimising the a priori assumptions. `Special' systems derived in this way are able to reveal important mechanisms absent in most other systems (Hogeweg, 1998).
The model developed by Hogeweg (2000b,a) evolves highly non-generic and complex gene networks, which, due to the unfolding and feedback to multiple intermediate levels and different time-scales, are difficult to comprehend in terms of the observed morphogenesis. Nevertheless, we can still study this model system relatively easily, and uncover general principles that also apply to the complex gene networks found in nature.
Although this thesis ends with the culmination of Dictyostelium discoideum morphogenesis, there are still a number of gaps in our knowledge. Our model also points to directions for interesting further research. As indicated before, we have not yet shown how the transition takes place from the mound stage to the slug stage, or from the slug stage to the culmination stage. We have worked on this problem, but we still do not completely understand the mechanisms underlying these transitions and the way they are timed. As is known from experiments, NH3 again plays an important role here, and hence we have focused mainly on this signalling molecule.
Another problem is related to phototaxis. Fisher (1997) tried to explain bi-directional phototaxis of certain mutant slime mould slugs as an actively turning towards and away from the light, so as to correct the deviation from the preferred direction of motion. We, however, found that the bi-directionality occurred simply as a side-effect of the turning mechanism, when the slug is too sensitive to NH3. Thus, we have found a mechanism which brings together multiple aspects of the phototaxis; an extra validation of the model. A recent paper by Miura & Siegert (2000), however, has shown that, with respect to phototaxis, life is not so simple. They showed that a sudden increase in light intensity leads to a transient appearance of new cAMP waves, in between those that are already propagating. Moreover, provided there are no spiral or target waves, new target waves can appear. These effects last during 3-4 waves. The experiments show that light does indeed influence the cAMP waves; at first glance, however, in the opposite way to the way we assumed in chapter 4. However, to explain phototaxis, one cannot ignore, for example, the experimentally established lens-effect. Hence, these experiments have to be interpreted within the framework of our model, if we want to understand how the observed effects can contribute to the many features related to phototaxis.
Because multiple module simulations like the ones presented in this thesis consume a lot of computer time, all simulations were done using a two-dimensional (2D) grid (except for a number of small test simulations). To be able to simulate the development in 2D, we modelled different stages using different viewpoints, i.e. either top-view (e.g. to model phototaxis), or side-view (e.g. to model the culmination).
Since the simulations were done in 2D, we still do not know whether our results are affected by this large simplification. We expect that all identified mechanisms will still hold in three dimensions (3D). Nevertheless, a number of behaviours are assumed to be purely 3D phenomena, such as tip formation, or the occurrence of a scroll wave in the anterior part, which breaks up in plane waves posteriorly. This could be important for the initiation and termination of the slug stage.
It would also be interesting to make the model for spiral breakup studied in part I into a 3D model. In most models for spiral breakup, the breakup is connected to instabilities in the propagation of wave-trains in excitable media. When this is simulated in a 3D homogeneous excitable medium, with an initial scroll wave, one can observe that the spiral breaks up in an identical way in every cross section : no additional 3D behaviour can be observed. This new mechanism of ours, however, has the potential to bring about breakups that are truly 3D, since wavefronts are laterally unstable in all directions. In 3D this may lead to interesting new dynamics.
In part II we have used a model in which the properties of the excitable medium depend on the history of the excitation pattern. Thus, our model could simply be viewed as an excitable medium in motion. This more general problem is closely connected with the questions that arise concerning the modelling of other kinds of excitable tissue. For example, in cardiac tissue the excitation wave also initiates a wave of motion (cardiac contraction), which can also affect excitation.
Finally, in our opinion it would be very challenging to start modelling embryogenesis itself, using the approach we have presented in this thesis, combined with the ever-increasing store of knowledge on gene networks, (limited) heredity and cell-to-cell signalling. This provides a framework in which cells can differentiate and generate shape simultaneously, and in which detailed experimental findings can be linked to the observed development of the whole organism.
The emergence of multicellular organisms is regarded as one of the major transitions in evolution. The cellular slime mould Dictyostelium discoideum has shown itself to be an excellent paradigm for the study of various aspects of multicellular development. It is a unique `model' organism, because it presents us within 48 hours with a complete transition to multicellularity, which is inevitably attended by multicellular coordination and morphogenesis. We have shown that the morphogenesis in the broadest sense of the word can emerge from the interactions and feedbacks between a few simple modules. We have presented a model consisting of a relatively small number of simple, well-defined and well understood building blocks, whose properties are in accordance with experimental findings, and which interact with each other only locally. This has given us an informative description of the Dictyostelium morphogenesis, from single cells all the way to the fruiting body. We have shown that in order to progress from (sub)cellular interactions towards a coordinated multicellular behaviour that is able to sense and respond to the environment in an appropriate way, the slime mould uses spatiotemporal patterns which are `larger than life'. Such patterns are formed by interacting processes, and are used for feedback between different levels of organisation. Finally, we have shown that even when complex dynamics emerge, it is still feasible to unravel the underlying mechanisms by using this multiple-scale modelling approach and looking at the intermediate scales.

